Close
Close

A History of Legs in 5 Objects
By Stacy Hackner, on 11 April 2017
 by Stacy Hackner
by Stacy Hackner
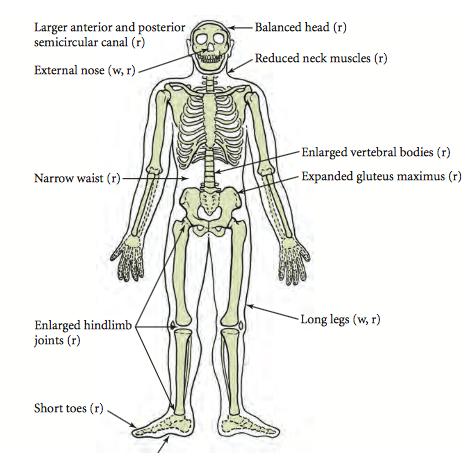
My research focuses on the tibia, the largest bone in the lower leg. You probably know it as the shin bone, or the one that makes frequent contact with your coffee table resulting in lots of yelling and hopping around; that’s why footballers often wear shinguards. The intense pain is because the front of the tibia is a sharp crest that sits directly beneath the skin. There are a lot of leg-related objects in UCL Museums, so here’s a whirlwind tour of a few of them!
One of the few places you can see a human tibia is the Petrie’s pot burial. This skeleton from the site of Badari in Egypt has rather long tibiae, indicating that the individual was quite tall. The last estimation of his height was made in 1985, probably using regression equations based on the lengths of the tibia and femur (thigh bone): these indicated that he was almost 2 meters tall. However, the equations used in the 80s were based on a paper from 1958, which used bone lengths from Americans who died in the Korean War. There are two problems that we now know of with this calculation: height is related to genetics and diet, and different populations have differing limb length ratios.

Pot burial from Hemamieh, near the village of Badari UC14856-8
The Americans born in the 1930s-40s had a vastly different diet from predynastic Egyptians, and the formulae were developed for (and thus work best when testing) white Americans. This is where limb length ratios come into play. Some people have short torsos and long legs, while others have long torsos and short legs. East Africans tend to have long legs and short torsos, and an equation developed for the inverse would result in a height much taller than he actually was! Another thing to notice is the cartilage around the knee joint. At this point in time, the Egyptians didn’t practice artificial mummification – but the dry conditions of the desert preserved some soft tissue in a process called natural mummification. Thus you can see the ligaments and muscles connecting the tibia to the patella (knee cap) and femur.
The Petrie also has a collection of ancient shoes and sandals. I think the sandals are fascinating because they show a design that has obviously been perfected: the flip flop. One of my favorites is an Amarna-period child’s woven reed sandal featuring two straps which meet at a toe thong. The flip flop is a utilitarian design, ideal for keeping the foot cool in the heat and protecting the sole of the foot from sharp objects and hot ground surfaces. These are actually some of the earliest attested flip flops in the world, making their appearance in the 18th Dynasty (around 1300 BCE).

An Egyptian flip-flop. UC769.
Another shoe, this time from the site of Hawara, is a closed-toe right leather shoe. Dating to the Roman period, this shows that flip flops were not the only kind of shoe worn in Egypt. This shoe has evidence of wear and even has some mud on the sole from the last time it was worn. This shoe could have been worn with knit wool socks, one of which has been preserved. However, the Petrie Collection’s sock has a separate big toe, potentially indicating that ancient Egyptians did not have a problem wearing socks and sandals together, a trend abhorrent to modern followers of fashion (except to fans of Birkenstocks).
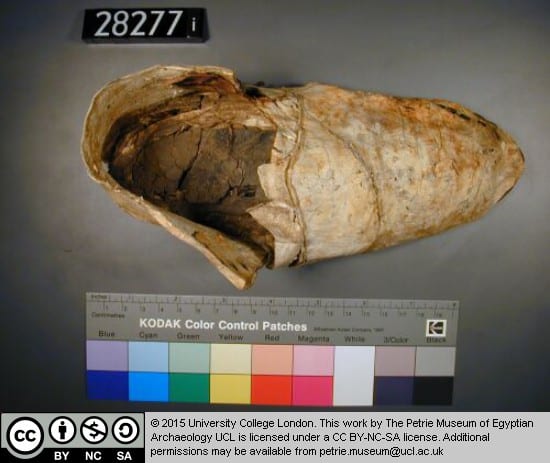
Ancient Egyptian shoe (UC28271).

Ancient Egyptian sock (UC16767).
The Grant Museum contains a huge number of legs, but only one set belonging to a human. For instructive purposes, I prefer to show visitors the tibiae of the tiger (Panthera tigris) on display in the southwest corner of the museum. These tibiae show a pronounced muscle attachment on the rear side where the soleus muscle connects to the bone. In bioarchaeology, we score this attachment on a scale of 1-5, where 5 indicates a really robust attachment. The more robust – attachment, the bigger the muscle; this means that either the individual had more testosterone, which increases muscle size, or they performed a large amount of activity using that muscle. (We wouldn’t score this one because it doesn’t belong to a human.) In humans, this could be walking, running, jumping, or squatting. Practice doing some of these to increase your soleal line attachment site!
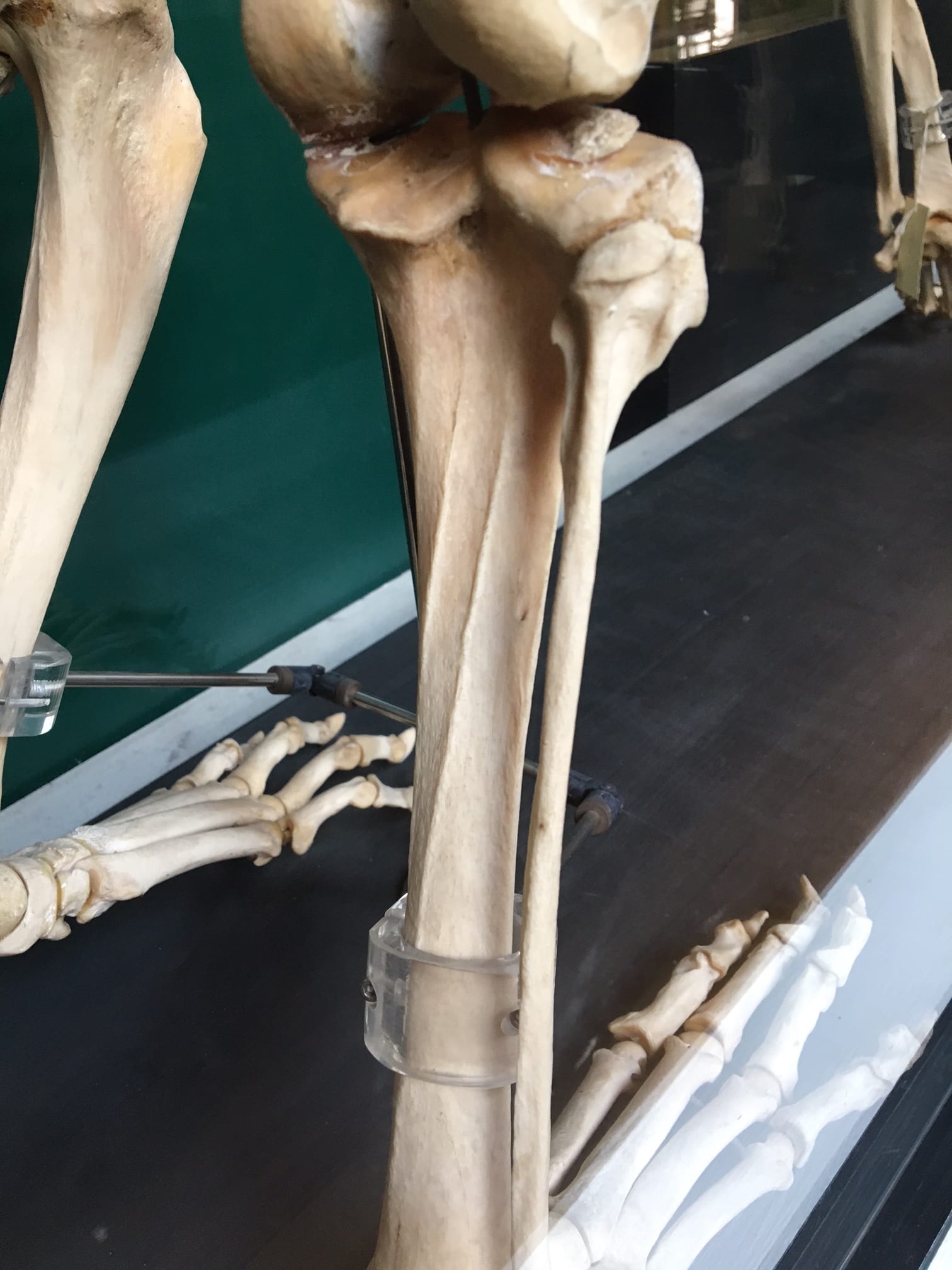
The posterior tibia of a tiger.
Moving to the Art Museum, we can see legs from an aesthetic rather than practical perspective. A statue featuring an interesting leg posture the legs is “Spinario or Boy With Thorn”, a bronze statue produced by Sabatino de Angelis & Fils of Naples in the 19th century. It is a copy of a famous Greco-Roman bronze, one of very few that has not been lost (bronze was frequently melted down and reused). The position of the boy is rather interesting: he is seated with one foot on the ground and the opposite foot on his knee as he examines his sole to remove a thorn. This is a very human position, and shows the versatility of the joints of the hip, knee, and ankle. The hip is adducted and outwardly rotated, the knee is flexed, and the ankle is everted. It’s rare for the leg to be shown in such a bent position in art, as statues usually depict humans standing or walking.

Spinario, or Boy With Thorn.
Bipedalism, or walking on two legs, is one of the traits we associate with being human. It’s rare in the animal world. Hopefully next time you look at a statue, slip on your flip flops, or go for a jog, you’ll think of all the work your tibiae are doing for you – and keep them out of the way of the coffee table.
(OK, I know that was six objects… but imagine the sock inside the shoe!)